목차
일반적인 유전자의 구조와 발현
- 유전자는 intron과 exon으로 구성되어 있다.
- Exon은 성숙한 mrna로 전사되는 염기서열이며 intron은 성숙한 mrna로 되기 전 제거되는 염기서열이다.
- 유전자의 5’ 말단에는 leader sequence가 존재하며 이것은 약 10개의 소수성 아미노산으로 이루어진 signal peptide를 단백질의 N 말단에 코딩한다.
- Leader Sequence 앞 부분에는 전사인자와 전사를 위한 RNA polymerase가 결합하여 전사를 조절하는 부위가 존재한다.
- 전사 이후 5’ 말단의 methylguanine에 모자(cap)이 그리고 3’ 말단의 꼬리부분에 약 205개의 adenine이 붙는 poly A tail이 생긴다.
- 스플라이싱 : 이 과정을 통해 전사체의 intron이 잘려나가고 전체 exon부분이 연결된 성숙한 mrna가 생성된다.
- mRNA는 핵을 떠나 ribosome에서 단백질로 translation 된다.
- 엑손은 일반적으로 세포외도메인, 막관통도메인, 세포질도메인의 각 부분을 발현할 수 있다.
- B세포 표면에 발현되어 있는 막 결합 Ig도 일반적인 세포표면 분자와 유사

Ig 사슬 합성을 위해 유전자 수준에서 일어나는 일
- 경사슬 유전자 (κ와λ)는 크게 두 부위로 나뉘는데 하나는 가변부위VL 로서 경사슬 N 말단의 약 108개의 아미노산으로 구성된다 또 하나는 불변부위CL이다.
일반적인 특징
- 경사슬의 가변부위를 코딩하는 2개의 서로 다른 유전자, 가변 분절 (V)
- 경사슬의 불변부위를 코딩하는 유전자, 불변 분절 (C)
- 가변부위의 코딩은 V유전자와 J유전자를 잘라 유전체에서 V(D)J recombinase로 알려진 효소 복합체에 의해 연결된다. 연결된 VJ 유전자 분절은 C 유전자와 함께 완전한 경사슬을 합성한다. V(D)J 유전자의 조합은 항체의 가변부위에 다양성을 형성 시킨다.
- 재조합 효소는 소위 V(D)J라고도 하는데 이 이유는 중사슬은 V, J 분절뿐만 아니라 D 분절도 사용하고 같은 재조합 효소가 경사슬과 중사슬 DNA의 자름과 연결에 관여하고 있기 때문이다.
Κ 사슬 합성
- 배선 (germline)의 염색체 2번에서 발견되는 κ 사슬은 40여개의 Vκ 유전자 분절이 선형으로 배열되어 있으며 intron에 의해 분절이 분리되어 있다 또한, 5개의 서로 다른 Jκ 분절이 Vκ아래에 늘어서 있고 하나의 Cκ가 Jκ 유전자의아래 부분에 위치한다.
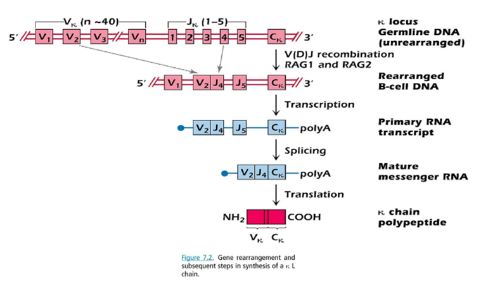
- 각각의 Vκ유전자 분절은 κ가변부위의 95개 아미노 말단 아미노산을 코딩할 수 있다. 각 Jκ분절은 나머지 13개의 아미노사(96-108)을 코딩한다
- V와 J의 유전자 선택 기작은 아직 불분명하지만 아마도 무작위적인 과정일 것으로 추측된다
- 이러한 유전자 재배열은 재조합효소 복합체에 의해 매개되는데 이들은유전자 분절을 자르고 연결한다
- 대표적인 효소는 RAG-1 (recombination activating gene I)과 RAG-2가발달 과정에 있는 림프구에서 발현되어 Ig와 T 세포 수용체 DNA를 자르는첫 단계에 요구되며 T 세포와 B 세포의 발달에 중요하다
- V(D)J 재조합효소는 V와 J 유전자 분절의 끝에 위치한 재조합 인식 염기서열 (recombination recognition sequences)을 인식한다
- 재조합 효소는 V2와 J4 유전자 분절을 연결시키고 사이에 존재하는 유전자는 고리 구조를 형성하여 잘려나간 후 분해된다
- B 세포에서 DNA가 재배열된 후 intron을 포함한 1차 전사체가 만들어진다. 이 전사체는 splicing에 의해 intron이 제거되고 exon이 결합되어성숙한 mRNA를 형성한다.

λ 사슬 합성
- 인간의 22번 염색체에서 발견되며, λ유전자의 재배열은 2개의 κ유전자좌에서의 재배열이 성공적으로 수행되지 않았을 때 일어난다
- λ사슬의 재배열 방법도 κ사슬의 재배열과 유사하게 V와 J 분절을 이용하고 V(D)J 재조합 효소에 의해 매개된다
- 30개의 Vλ유전자 분절 중 하나가 4개의 Jλ유전자 분절 중 하나와 연결되어 splicing이 일어나며 유전자 재배열이 일어난다.
중사슬 유전자의 구성과 재배열
- 사람의 H 유전자는 14번 염색체에 존재하며 각각의 경사슬과 유전자좌와 다른 위치에 있다.
- 차이점 : 중사슬의 가변영역을 코딩하는 유전자는 Vh, Dh, Jh의 3개의 유전자 분절로 구성된다. 따라서 V와 J와 함께 다양성 D 분절이 가변영역에 포함된다.
- 약 40개의 Vh, 약 25개의 Dh, 6개의 Jh로 구성된다.
- 특히, D와 J 분절은 중사슬의 CDR3 (complementarity determining region 3)라 불리는 아미노산을 코딩한다.
- V 부위 유전자는 2개의 CDR1, CDR2를 코딩한다. CDR1,2,3 는 가변부위에 존재.
- 인간의 중사슬은 3’말단에 밀집된 9개의 C 부위 유전자를 갖는다
- C 부위는 항체의 클래스 (IgM, IgD, IgG, IgE, IgA)를 결정하여 결과적으로 항체의 기능을 결정한다
- 경사슬과 같이 V(D)J 재조합 효소에 의해 재배열이 작동된다
- B 세포의 초기에는 두단계에 걸쳐 배선 DNA 재배열이 일어난다 첫째는 D와 J 분절의 결합이며, 두번째는 하나의 V 분절을 DJ 단위체에재조합하는 것으로 중사슬의 항원 특이성이 정해진다
- 성숙 B 세포는 재배열된 DNA에서 alternative splicing이라는 방법에의해 두개의 다른 성숙 mRNA를 생성한다. 번역과정을 거쳐 경사슬과조합되어 항원 특이성을 갖는 IgD와 IgM을 발현한다

클래스 또는 개별형 전환
- 하나의 B 세포는 VJ(경사슬)와 VDJ(중사슬) 재배열에 의해 고정되는 단일특이성을 나타내는 항체를 만든다
- IgM+ IgD+ 성숙 B세포가 항원 자극과 동시에 T 세포의 신호가 있을 때, 자극되면 이는 클래스 전환 재조합 (class switching recombination)을통해 class 또는 isotype 전환이 일어난다.
- 재배열된 VDJ가 중사슬의 불변부위의 다른 C 유전자와 결합한다. 이때 각 C 유전자의 변화는 5’말단에 존재하는 switch region (S) 사이에서 일어나며사용되지 않은 S 부위를 포함한 C 부위 DNA는 제거된다.
- 이후 재배열된 1차 전사체는 splicing에 의해 intron이 잘려 나간 후 mRNA를 생성하고 최종적으로 B 세포는 IgG, IgE, IgA와 같은 다른 class의항체분자를 합성한다
- Class 전환은 항체의 기능을 변화시키나 항원 특이성은 변환시키지는 않는다
- 클래스 전환 재조합은 일반적으로 2차 림프조직의 배중심(germinal center)에서 일어난다
- 특히, 이 과정은 BCR을 통한 항원 자극, B 세포와 T 세포 간의 상호작용, T 세포 유래의 cytokine을 요구한다.
- 또한, 2차 림프조직의 배중심에서 활성화된 B 세포가 발현하는 AID (activation-induced cytidine deaminase)를 필요로 하기도 한다.
- AID는 활성화된 B 세포에서 cytidine으로부터 아미노기를 제거하여 uridine으로 변환시킨다. DNA를 합성하는 과정에서 mutation이 일어남 -> 다양한 형태의 항체 생성
- 이로 인해 DNA에서 C:G가 아닌 U:G가 형성되면 다른 효소 작용에 의해 잘려지게 된다. 그 결과 Ig의 가변부위 유전자 DNA상에 변이를 유발하여항체의 다양성 형성에 중요한 역할을 한다.

항체의 다양성 형성
배선에 있는 다중의 V 유전자의 존재
- 배선 상에 존재하는 다수의 V 유전자는 항체 다양성의 근간으로 생성될 수 있는 최소한의 항체 수를 나타냄.
VJ와 VDJ 조합적 결합 => 다양한 항체 형성
- 경사슬은 VJ 결합을, 중사슬은 VDJ 결합을 할 수 있다. 이들은 무작위적 조합을 통해 다양한형태의 경사슬과 중사슬을 형성할 수 있다.
- 예를 들어, 40여개의 Vκ와 5개의 Jκ가 무작위적으로 조합한다고 가정--> 200개의 κ사슬을 형성, 30개의 V λ rhk 4개의 J λ은 120개의 λ사슬을 형성, 40개 V 유전자, 25개 D 유전자, 6개의 J 유전자는 6,000가지 중사슬을 형성
중사슬과 경사슬의 무작위적 조합 (random assortment)
- κ사슬을 포함한 Ig 분자는 200 x 6,000으로 200 * 10^6개, , λ사슬을 포함한 Ig 분자는 12 x 6,000으로 0.72 *10^6개의 분자를 형성할 수 있다.
연결 부위 다양성
- V(D)J 재조합으로 DNA 가닥은 잘리고 다시 연결된다 그러나 V, D, J 분절의 연결이 항상 정확하지는 않다 즉 연결되는 과정에서 몇 개의 nucleotide가 결손 혹은 첨가되기도 한다.
- 이러한 부정확한 DNA 재조합은 Ig의 항원과의 상보성이 결정되는 초가변부위에서 발생하기때문에 항원 결합 부위
체세포 고도변이
- 이 변이는 항체의 중사슬 혹은 경사슬의 VJ와 VDJ 재조합체에서 point mutation이 일어나서 아미노산 서열이 변하는 현상이다
- 정상적인 돌연변이 비율보다 적어도 10,000배 이상 더 높게 발생한다
- 일반적으로 항원에 대한 1차반응에서는 낮은 친화도를 갖는 항체가 생성되나, 2차 림프조직의 배중심에서 항원 자극에 의해 B 세포가 발달하면 항원에 대한 친화력이 증가한항체가 생성되며 이를 친화력 성숙 (affinity maturation)이라고 한다.
체세포 유전자 변환
- 조류와 토끼에서는 사람과는 다른 유니크한 다양성 생성 기작이 밝혀짐 => Pseudogenes
- 이 현상은 유전자 간 비상호적 염기서열 교환에 기초한다. 공여 유전자의 일부가 수여 유전자로 복제되어 수여 유전자가 변형된다
- 이러한 현상을 일으키는 기작은 정확히 밝혀져 있지 않다
- VH 위쪽에 돌연변이로 인해 polypeptide로 합성되지 못하는 약 20개의 pseudogene이 존재하며 이들의 짧은 염기 서열이 VDJ 유전자에 삽입되어 다양화된 가변부위 유전자가 생성된다
